Assessment |
Biopsychology |
Comparative |
Cognitive |
Developmental |
Language |
Individual differences |
Personality |
Philosophy |
Social |
Methods |
Statistics |
Clinical |
Educational |
Industrial |
Professional items |
World psychology |
Biological: Behavioural genetics · Evolutionary psychology · Neuroanatomy · Neurochemistry · Neuroendocrinology · Neuroscience · Psychoneuroimmunology · Physiological Psychology · Psychopharmacology (Index, Outline)
See:Visual cortex for overview of complete visual area of the brain
Primary visual cortex (V1)[]
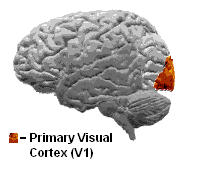
The primary visual cortex is the best studied visual area in the brain. Like that of all mammals studied, it is located in the posterior pole of the occipital cortex (the occipital cortex is responsible for processing visual stimuli). It is the simplest, earliest cortical visual area. It is highly specialized for processing information about static and moving objects and is excellent in pattern recognition.
The functionally defined primary visual cortex is approximately equivalent to the anatomically defined striate cortex. The name "striate cortex" is derived from the stria of Gennari, a distinctive stripe visible to the naked eye that represents myelinated axons from the lateral geniculate body terminating in layer 4 of the gray matter.
The primary visual cortex is divided into six functionally distinct layers, labelled 1 through 6. Layer 4, which receives most visual input from the lateral geniculate nucleus (LGN), is further divided into 4 layers, labelled 4A, 4B, 4Cα, and 4Cβ. Sublamina 4Cα receives most magnocellular input from the LGN, while layer 4Cβ receives input from parvocellular pathways.
Function[]
V1 has a very well-defined map of the spatial information in vision. For example, in humans the upper bank of the calcarine sulcus responds strongly to the lower half of visual field (below the center), and the lower bank of the calcarine to the upper half of visual field. Conceptually, this retinotopy mapping is a transformation of the visual image from retina to V1. The correspondence between a given location in V1 and in the subjective visual field is very precise: even the blind spots are mapped into V1. Evolutionarily, this correspondence is very basic and found in most animals that possess a V1. In human and animals with a fovea in the retina, a large portion of V1 is mapped to the small, central portion of visual field, a phenomenon known as cortical magnification. Perhaps for the purpose of accurate spatial encoding, neurons in V1 have the smallest receptive field size of any visual cortex regions.
The tuning properties of V1 neurons (what the neurons respond to) differ greatly over time. Early in time (40 ms and further) individual V1 neurons have strong tuning to a small set of stimuli. That is, the neuronal responses can discriminate small changes in visual orientations, spatial frequencies and colors. Furthermore, individual V1 neurons in human and animals with binocular vision have ocular dominance, namely tuning to one of the two eyes. In V1, and primary sensory cortex in general, neurons with similar tuning properties tend to cluster together as cortical columns. David Hubel and Torsten Wiesel proposed the classic ice-cube organization model of cortical columns for two tuning properties: ocular dominance and orientation. However, this model cannot accommodate the color, spatial frequency and many other features to which neurons are tuned. The exact organization of all these cortical columns within V1 remains a hot topic of current research.
Current consensus seems to be that early responses of V1 neurons consists of tiled sets of selective spatiotemporal filters. In the spatial domain, the functioning of V1 can be thought of as similar to many spatially local, complex Fourier transforms. Theoretically, these filters together can carry out neuronal processing of spatial frequency, orientation, motion, direction, speed (thus temporal frequency), and many other spatiotemporal features. Experiments of V1 neurons substantiate these theories, but also raise new questions.
Later in time (after 100 ms) neurons in V1 are also sensitive to the more global organisation of the scene (Lamme & Roelfsema, 2000). These response properties probably stem from recurrent processing (the influence of higher-tier cortical areas on lower-tier cortical areas) and lateral connections from pyramidal neurons (Hupe et al 1998).
The visual information relayed to V1 is not coded in terms of spatial (or optical) imagery, but rather as the local contrast. As an example, for an image comprising half side black and half side white, the divide line between black and white has strongest local contrast and is encoded, while few neurons code the brightness information (black or white per se). As information is further relayed to subsequent visual areas, it is coded as increasingly non-local frequency/phase signals. Importantly, at these early stages of cortical visual processing, spatial location of visual information is well preserved amid the local contrast encoding.
Current research[]
Research on the primary visual cortex can involve recording action potentials from electrodes within the brain of cats, ferrets, mice, or monkeys, or through recording intrinsic optical signals from animals or fMRI signals from human and monkey V1.
One recent discovery about V1 is that signals measured by fMRI show very large attentional modulation. This result strongly contrasts with macaque physiology research showing very small changes (or no changes) in firing associated with attentional modulation. Research with the macaque monkey is usually performed by measuring spiking activity from single neurons. The neural basis of the fMRI signal on the other hand is mostly related to post synaptic potentiation (PSP). This difference therefore does not necessarily indicate a difference between macaque and human physiology.
Other current work on V1 seeks to fully characterize its tuning properties, and to use it as a model area for the canonical cortical circuit.
Lesions to primary visual cortex usually lead to a scotoma, or hole in the visual field. Interestingly, patients with scotomas are often able to make use of visual information presented to their scotomas, despite being unable to consciously perceive it. This phenomenon, called blindsight, is widely studied by scientists interested in the neural correlate of consciousness.
See also[]
References & Bibliography[]
Key texts[]
Books[]
- Peters, Alan (ed), and Kathleen S. Rockland (ed). (1994). Cerebral Cortex: Primary Visual Cortex in Primates v. 10 (Cerebral Cortex). Kluwer Academic / Plenum Publishers
- Peters, Alan (ed) and Bertram Payne (ed). (2001). The Cat Primary Visual Cortex Academic Press.
Papers[]
Additional material[]
Books[]
Papers[]
External links[]
| Sensory system - Visual system - edit |
|---|
| Eye | Optic nerve | Optic chiasm | Optic tract | Lateral geniculate nucleus | Optic radiation | Visual cortex |